
1. Introduction
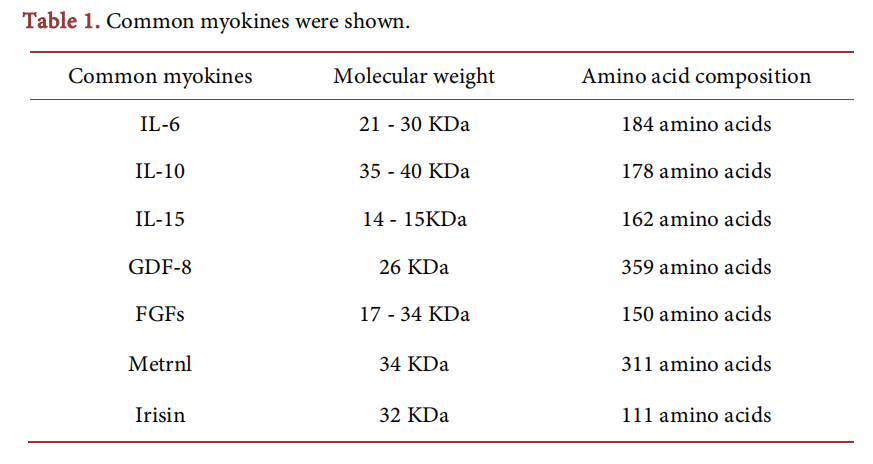
Diabetes mellitus is a metabolic disease caused by absolute or relative insufficiency of insulin secretion and is mainly manifested as hyperglycemia. Statistics from the Worth Health Organization demonstrate that about 0.46 billion people were attacked by diabetes mellitus worldwide in 2021 [1] [2]. Diabetes mellitus is mainly divided into type 1 diabetes mellitus (T1DM) and type 2 diabetes mellitus (T2DM) as well as gestational diabetes mellitus (GDM), monogenic diabetic syndrome, and drug- or chemical-induced diabetes mellitus [3]. T2DM is most common in clinic, accounting for about 90% of all diabetes mellitus and mostly occurs in middle-aged and old people [4]. T2DM, the fifth cause of death world-wide [5], often occurred in the background of insulin resistance. When the function of β cells in the pancreatic islet fails to compensate for insulin resistance, hyperglycemia and impaired glucose tolerance eventually lead to T2DM. With the application of exercise therapy in diabetes mellitus and other metabolic diseases [6], the indispensable roles of skeletal muscles have attracted wide attention. Skeletal muscles are important organs in physical exercises, posture maintenance and energy metabolism regulation. They are important endocrine organs, secreting various myokines through autocrine or paracrine function [7]. Skeletal muscles participate in energy metabolism, insulin resistance, glucolipide metabolism and other processes [7]. Interleukin (IL)-6, the first named myokine, is positively correlated with the incidence of diabetes mellitus [8]. Other myokines including IL-10, IL-15, growth differentiation factor (GDF)-8, fibroblast growth factor (FGF), meteorin-like (metrnl), and irisin were discovered subsequently [9] [10]. The levels of myokines in surm will change during insulin resistance and exercise therapy for diabetes mellitus, which can be used as a target for diabetes therapy and prognosis. In this work, the development of common myokines (Table 1) and effects on the occurrence and development of diabetes mellitus as well as the roles of myokines in diabetes mellitus are reviewed. This work emphasizes the role of myokines, and provides evidence for the prevention, early diagnosis and treatment of diabetes mellitus.
1.1. IL-6
IL-6, the first discovered myokine, has a molecular weight of 21 - 30 KD and is composed of 184 amino acids. Initially, IL-6 was thought as an inflammatory factor. When the body is under the inflammatory status, monocytes and macrophages rapidly secrete IL-6 and thereby induce C-reactive protein and procalcitonin to participate in the inflammation process. Previous evidence has shown that in addition to mediating inflammation, IL-6 also regulates glucose homeostasis and lipid metabolism [11].
A meta-analysis demonstrates that IL-6 disturbs insulin signals and damages the functions of islet β cells [8], thereby participating in the occurrence of T2DM, and IL-6 level is positively correlated with the incidence of T2DM [12]. In addition, IL-6 plays a dual role in maintaining blood glucose stability. IL-6 is a beneficial glucose metabolism regulatory factor in the healthy body, but may increase the severity of insulin resistance in obese or inflammatory patients. Kurauti et al. found that IL-6 may enhance insulin resistance by increasing the ratio of cyclic adenosine phosphate (cAMP) and AMP/ATP in rat skeletal muscle to activate 5'-adenosine monophosphate activated protein kinase (AMPK) to enhance the expression and activity of insulin degrading enzymes in liver and skeletal muscle, avoiding the sudden drop of blood glucose level during exercising [13]. Other studies have found [5] that IL-6 not only improves glucose uptake by stimulating AMPK and phosphatidylinositol 3 kinase (PI3K) activity, but also impacts insulin in a dependent manner by C-Jun amino-terminal kinase (JNK) [14]. Additionally, the IL-6 level is also associated with the subclinical inflammation and obesity of T2DM, indicating weight control may be a measure to prevent inflammation in T2DM patients.
The coronary artery disease (CAD) is a major cause for the morbidity and mortality of T2DM patients. There is increasing evidence that chronic vascular inflammation is driven by inflammatory factors and can mediate vascular complications of T2DM. Thus, it is significant to study the increase of inflammatory cytokine IL-6 in T2DM patients. Recent studies have suggested that, IL-6 is related to left ventricle dysfunctions and can be one of the methods to detect left ventricle dysfunctions in T2DM patients [15]. Elevated IL-6 levels can interfere with glucose metabolism and lead to insulin resistance, which together with tumor necrosis factor (TNF)-α and IL-1β can induce chronic inflammation and promote the occurrence of CAD in diabetic patients. Adela et al. found that IL-6 was elevated in both CAD and T2DM-CAD groups, especially in THE T2DM-CAD group, by studying the serum protein characteristics of CAD in Indian T2DM patients [16]. Moreover, high blood glucose level can lead to inflammation of vascular endothelial cell (EC) and migration and proliferation of smooth muscle cells (SMC) through activating the JAK-STAT signal to stimulate IL-6 release [17]. As a result, high IL-6 level is associated with the vascular EC inflammation and uncontrollable SMC migration/proliferation in T2DM patients.
1.2. IL-10
IL-10, also known as cytokine synthesis inhibitor, is a multifunctional inflammatory suppressor that plays an immunological role in many cell types. IL-10 is mainly produced by Th2 cells and mononuclear macrophages and can inhibit the production of cytokines by activated T cells (especially the production of IL-2 and IFN-γ by Th1 cells), which inhibte cell immunity response. In addition to the important regulating effects in immune responses in vivo, there is growing evidence that IL-10 is involved in type 2 diabetes [18] [19] [20].
At locus 1082 of transcription initiation loci, the existence of G allelic gene is related to the high expression of IL-10. IL-10 inhibits the production of in-flammatory factors by inhibiting T cells, monocytes and macrophages, and the IL10-1082G/G genotype can down-regulate the immune response [21] [22]. A meta-analysis involving 4250 subjects demonstrates that the IL10-1082G allelic gene is linked to a higher risk of T2DM [23]. An analysis involving 490 T2DM patients and 349 Italian Caucasians shows that the subjects with IL10-597A/ -824T/-1087A negativity are more prone to the main vascular damages dominated by T2DM [24]. The clinical significance of IL-10 level in insulin resistance has also been confirmed, and IL-10 level is positively correlated with insulin sensitivity [25]. Meanwhile, Zeng Jing et al. showed that IL-10 was negatively correlated with the severity of diabetic peripheral neuropathy [26].
1.3. IL-15
IL-15, which belongs to the 4α spiral cytokine family, is widely expressed in various cells and tissues, and was involved in 2007 into the group of myokines [27]. Though the mRNA that regulates IL-15 has been discovered in various cells and tissues, the IL-15 protein is only expressed in specific cells, such as skeletal muscle cells [28]. And the IL-15 in blood plasma is mainly secreted by skeletal muscles [29]. Studies have found that IL-15 levels are slightly increased or unchanged after acute exercise [30]. However, the important roles of IL-15 in regulating glucose metabolism, promoting skeletal muscle glucose uptake and immune regulation cannot be ignored.
T1DM is mainly caused by T and B lymphocytes synergistic action to attack islet cells resulting in absolute insulin deficiency [31]. In the normal environment in vivo, CD8+ Tregs can terminate the interaction of T-B lymphocytes and thereby protect islet cells from attacks. Stocks et al. found that in NOD mice, a large dose of IL-15 could increase the number and activity of CD8+ Tregs cells and inhibit the function of T-B cells, thereby reduce blood glucose level and relieve T1DM [32], which provides new clue for T1DM treatment. Insulin resistance is a major indicator in the pathogenesis of T2DM. IL-15 can bind with its receptor (IL-15R) to form a membrane-binding IL-15-IL-15R composite, which will activate the phosphorylated JAK (Janus kinase)/STAT (signal transducer and activator of transcription proteins) Signaling pathways, initiating signal transduction, inhibiting gluconeogenesis and decreasing insulin sensitivity and blood sugar level in vivo [33]. Study has confirmed that high-dose IL-15 can cause metabolic adaptation, improve insulin sensitivity and decrease the blood glucose level and insulin resistance of diabetic patients [34]. High-fat diet mice are often accompanied with the rise of blood glucose levels, which the muscular IL-15 level rises and blood glucose level drops through running on the wheel [35]. A 6-year prospective study involving 6537 non-diabetic children in 8 European countries shows that the IL-15 level is negatively correlated with the insulin resistance index, indicating the IL-15 level can be a target of prevention or treatment at early stage of diabetes mellitus [32].
Concluding, IL-15 may play an critica role in the pathogenesis, monitoring and intervention of T1DM, T2DM and obesity. However, so far, there are few studies on IL-15 in human body, and its definite mechanism is still unclear,
1.4. GDF-8
GDF-8, belongs to the transforming growth factor β superfamily, is a key regulator of skeletal muscle mass. In 1997, McPherron et al. discovered GDF-8 in mouse muscle tissues by using PCR (polymerase chain reaction), and found the skeletal muscle in GDF-8-deficient mice was 2 to 3 times that in wild-type mice [36]. After, GDF-8 was also known as MSTN (myostatin) because of its inhibitory effect on skeletal muscle growth. The existing research implied that GDF-8 is involved in the pathophysiological processes of cardiovascular diseases, kidney diseases, obesity and diabetes. which needs to be further studied by expanding samples.
Amor M et al. found that GDF-8 is upregulated in the serum of obese patients and is positively correlated with the insulin resistance index, while negatively with the insulin sensitivity index [37]. Insulin resistance is a key factor for the occurrence of T2DM. When the function of pancreatic β cells cannot compensate for insulin resistance, abnormal rise of blood glucose level and impaired fasting glucose will occur, which finally lead to T2DM [38]. A cross-sectional study involving 264 T2DM patients indicates that the level of GDF-8 is positively correlated with retinopathy [39] and affects the progression and prognosis of diabetes. Downregulation of GDF-8 in T2DM patients can active the insulin signaling pathway, antagonizes the negative effects of on muscle mass, muscle strength and insulin signaling pathway partly in T2DM mice par, and thereby improve glucose homeostasis and relieve insulin resistance [40]. In DM mice, metformin can significantly regulate GDF-8-mediated muscle dysfunction in skeletal muscle cells through AMPK-FOXO3A-HDAC6 pathway [41], providing a new idea for DM drug diabetes mellitus therapeutic development. Dapagliflozin, a T2DM therapeutic drug, can significantly downregulate the GDF-8 level, maintain skeletal muscle mass and relieve diabetes-related muscle tissue loss [42]. Emerging diabetes exercise therapy showed excellent effect on diabetic rats [43]. Clinical research indicate that aerobic training combined with resistance training can increase GDF-8 level of T2DM patients, and relieve the diabetesinduced amyotrophia symptoms [44], which may be related to the high miR-23a/27a expression in muscle tissues , but the definite mechanism needs further research. GDM attacks women with normal normal glucose metabolism or potential glucose tolerance reduction during pre-pregnant and is occurring or diagnosed during gestation. It is adverse to fetal growth and development. Early maternal serum marker GDF-8 increased in patients with GDM, hlghly GDF-8 has predictive value for the occurrence of GDM at late trimester of pregnancy [45].T1DM, also called insulin-dependent diabetes mellitus, is related to the immunity-mediated destruction of β cells in late trimester of pregnancy [46]. Clinical research implies that the serum GDF-8 level of T1DM children is significantly higher than healthy children at the same age and BMI (body mass index), which may be associated with impaired glucose tolerance [2]. Interestingly, Dial A G et al. found the GDF-8 level was not fully related to clinical indices. The skeletal muscles of pigs can be enhanced and even the glucose ingestion in T1DM can be improved by controlling GDF-8 in vivo [47]. Hence, the function of GDF-8 in T1DM may involve complex regulatory mechanisms, and exploring the mechanism of GDF-8 upregulation and its correlation with insulin dose in T1DM is extremely important.
1.5. FGF
FGFs are a structure-related polypeptide family and also called heparin-binding growth factors. FGFs in the form of paracrine or endocrine can mediate body development, tissue homeostasis or restoration, and relevant metabolic activities [48]. Recent research shows that FGF-1, FGF-4, FGF-19, and FGF-21 can significantly act on diabetes mellitus and can be used as targets for early diagnosis,prevention and treatment of diabetes mellitus.
FGF1, the first discovered member in this family, is a strong angiogenesis factor and pro-mitogen that is a signaling protein encoded by the FGF1 gene. In vitro research demonstrates that all the subtypes (FGFR1, FGFR2, FGFR3, FGFR4) of the FGF receptor as well as FGF1 are expressed at the mRNA level in the insular tissues and pancreas β cell INS-1E cell line of rats, and FGF1 can enhance the proliferation of INS-1E cells and improve the activity of insular cells [49]. After the stimulus of islet cells by FGF1, the phosphorylation levels of extracellular regulated protein kinases (ERK1/2) are significantly increased, and blockage of ERK1/2 can destroy the exciting effect of FGF1 on islet cell proliferation, indicating ERK-related pathways are involved in the islet cell proliferation and regulation by FGF1 [50]. Experiments on T2DM and T1DM mice demonstrate that FGF1 can improve the rising blood sugar level in T2DM mice, but not in T1DM mice, suggesting the reduction of blood sugar level by FGF1 must be achieved with the presence of islet cells. This finding offers some theoretical basis for FGF1 to directly stimulate islet cells [51].
FGF4 plays an indispensable role in embryonic development, including implantation, morphogenesis, and organogenesis. FGF4 was found to be an effective anti-hyperglycemic factor with hypoglycemic effects comparable to FGF1[52]. This study also validates that the endocrine FGF21 and FGF19 regulate hypoglycemic activities mainly in the liver, but not in muscles. Hence, FGF19 and FGF21 cannot stimulate the glucose ingestion by skeletal muscles, which explains the fact that FGF secretion in skeletal muscle is worse than paracellular secretion.
FGF19 was first discovered in the brain at the human fetal development stage, but was later found to be mainly expressed in the ileum. FGF19 not only plays a role as a physiological regulator of bile acid homeostasis, but also plays an insulin-like role in promoting glycogen synthesis and reducing blood glucose levels in diabetic mice. It is important to note that lateral ventricle injection of FGF19 can decrease the blood sugar levels of diabetic mice, without affecting insulin secretion or systemic insulin sensitivity, indicating FGF19 can reduce the blood sugar level in an insulin-independent way [53] [54].
FGF21 is mainly expressed in the liver and slightly expressed in extrahepatic tissues, such as white and brown fat tissues, skeletal muscles, the heart, kidney and pancreas. FGF21 has an insulin-like effect and can improve the function of islet β cells by regulating the protein kinase B signaling pathway, thus regulating fat and glucose metabolism, which has varying degrees of elevation in patients with diabetes, coronary heart disease, obesity and metabolic syndrome [55] [56]. Hence, FGF-21 may also be a biomarker to indicate the higher risk of developing metabolic syndrome.
Altogether, the occurrence and development of diabetes mellitus are regulated by multiple factors in various signaling pathways. However, the heterogeneity among races, origin heterogeneity among tissues from different parts, and differences in microenvironment call for abundant experiments for data collection and research, which will theoretically underlie clinical applications.
1.6. Metrnl
Meteorin-like (Metrnl) is a newly-discovered myokine that is usually produced after exercises or cold exposure. Metrnl can inhibit the insular invasion of lymphocytes to improve insular inflammation and significantly delay the occurrence of diabetes mellitus. The peroxisome proliferators-activated receptor γ (PPARγ) is a key regulatory factor of insulin resistance and fat cell differentiation, and Metrnl can regulate the PPARγ pathway to affect insulin sensitivity. Thus, upregulation of Metrnl level may improve the autoimmune destruction of β cells and becomes a new strategy to increase blood glucose and insulin levels [57]. Wenchao Hu et al. verified that Metrnl can improve the functions of β cells by activating the WNT/β-catenin pathway, inhibiting β cell apoptosis and promoting β cell proliferation [58]. The existing research findings on the serum Metrnl levels in T2DM patients are not all consistent. Some studies show that the Metrnl levels of T2DM patients are lower than those in healthy controls [59] [60] [61], but Hye Soo Chung et al. found the Metrnl levels rose in T2DM patients [62]. Another study showed the Metrnl level in T2DM patients did not significantly change [63]. Thus, to study the relationship between Metrnl and T2DM, well-designed cohort research is needed, in which while all possible influence factors are excluded, the severity of diabetes mellitus must strictly scored, such as degree of blood glucose fluctuation, and complications.
The existing studies are mostly limited to the correlation between Metrnl level alone and diabetes mellitus, but there is no research on the correlation between diabetes mellitus and Metrnl when Metrnl is synergistic with other factors.
1.7. Irisin
Irisin, a novel myokine discovered in 2012, is composed of 111 amino acids and has a molecular weight of 32 KD. It is extensively distributed in organisms and can be released by skeletal muscle cells after exercise [23]. Research has confirmed that irisin plays a vital role in maintaining glucose and adipose homeostasis, and has potential therapeutic values for metabolic disorders such as type 2 diabetes and obesity.
Irisin is negatively correlated with fasting plasma glucose, insulin level, homeostasis model assessment-insulin resistance, retinol binding protein4, and hemoglobin in diabetic patients. The Irisin level is low in T2DM patients accompanied with vascular complications. The above findings indicate irisin has certain relieving effect on diabetes mellitus [64]. Tarboush et al. found that irisin level in plasma was not only correlated with DR (a diabetic retina lesion) stage of diabetic retinopathy, but also significantly different in irisin level between DR stages, which has great guiding value for studying irisin as a potential associated marker of diabetic complications [65]. Irisin can also relieve myocardial microvascular injuries, cell apoptosis, insulin resistance and blood sugar level rise through activating the ERK1/2/Nrf2/HO-1 pathway in diabetic patients [66]. Moreover, irisin can inhibit the autophagia of H9c2 cells and improve insulin resistance through activating the PI3K/Akt pathway [67]. In streptozotocin (STZ) induced diabetic mice, intraperitoneal injection of recombinant Irisin significantly reduced the daily water intake, food intake and blood glucose [68].
In all, irisin is closely related to sugar metabolism and has been shown to have a certain degree of therapeutic effect in alleviating diabetes in cells, animals and clinical experiments However, studies on the correlation between irisin and diabetes and metabolic syndrome also have different results One study shows that there is no significant correlation between serum irisin and glucose metabolism related indicators in T2DM patients [69]. As a result, the definite mechanism of irisin in diabetes mellitus calls for further research.
2. Conclusion and Prospects
Taken together, the myokines released during skeletal muscle contraction play important regulating roles involved in the development of diabetes, which may become potential targets for diagnosis, monitoring, prevention and treatment of diabetes mellitus. Diabetes patients lacking a specific muscle factor can be improved by targeting specific muscle factors, or as a new test indicator or a combination of indicators for the diagnosis of the disease. Moreover, exercise therapy can also be an effective way to treat diabetes or adjunctive therapy. Although myokines are closely related to the occurrence of diabetes mellitus, the underlying mechanism of how they play a role in the pathogenesis and target organ injury is still unclear and needs to be further explored. The results of cell, animal and even the clinical trial are a degree of heterogeneity or even fully opposite, which may be related to different sample sizes, selected population ethnicity, clinical characteristics of the disease, and measurement criteria and methods of myokine. Given the extensive applications of myokines in medicine and exercise medicine, exploring the role and mechanism of exercise or myokines in alleviating diabetes mellitus will be conducive to better playing the health promotion role of exercise.
Conflicts of Interest
The authors declare no conflicts of interest regarding the publication of this paper.
References
[1] Siehler, J., Blöchinger, A.K., Meier, M. and Lickert, H. (2021) Engineering Islets from Stem Cells for Advanced Therapies of Diabetes. Nature Reviews Drug Discovery, 20, 920-940. https://doi.org/10.1038/s41573-021-00262-w
[2] Dial, A.G., Monaco, C.M.F., Grafham, G.K., Romanova, N., Simpson, J.A., Tarnopolsky, M.A., Perry, C.G.R., Kalaitzoglou, E. and Hawke, T.J. (2020) Muscle and Serum Myostatin Expression in Type 1 Diabetes. Physiological Reports, 8, e14500. https://doi.org/10.14814/phy2.14500
[3] Lee, E.J., Ahmad, S.S., Lim, J.H., Ahmad, K., Shaikh, S., Lee, Y.S., Park, S.J., Jin, J.O., Lee, Y.H. and Choi, I. (2021) Interaction of Fibromodulin and Myostatin to Regulate Skeletal Muscle Aging: An Opposite Regulation in Muscle Aging, Diabetes, and Intracellular Lipid Accumulation. Cells, 10, Article No. 2083. https://doi.org/10.3390/cells10082083
[4] Galicia-Garcia, U., Benito-Vicente, A., Jebari, S., Larrea-Sebal, A., Siddiqi, H., Uribe, K.B., Ostolaza, H. and Martín, C. (2020) Pathophysiology of Type 2 Diabetes Mellitus. International Journal of Molecular Sciences, 21, Article No. 6275. https://doi.org/10.3390/ijms21176275
[5] Rizzo, M.R., Di Meo, I., Polito, R., Auriemma, M.C., Gambardella, A., di Mauro, G., Capuano, A. and Paolisso, G. (2022) Cognitive Impairment and Type 2 Diabetes Mellitus: Focus of SGLT2 Inhibitors Treatment. Pharmacological Research, 176, Article ID: 106062. https://doi.org/10.1016/j.phrs.2022.106062
[6] Daradics, Z., Crecan, C.M., Rus, M.A., Morar, I.A., Mircean, M.V., Cătoi, A.F., Cecan, A.D. and Cătoi, C. (2021) Obesity-Related Metabolic Dysfunction in Dairy Cows and Horses: Comparison to Human Metabolic Syndrome. Life (Basel), 11, Article No. 1406. https://doi.org/10.3390/life11121406
[7] Barbalho, S.M., Prado Neto, E.V., De Alvares Goulart, R., Bechara, M.D., Baisi Chagas, E.F., Audi, M., Guissoni Campos, L.M., Landgraf Guiger, E., Buchaim, R.L., Buchaim, D.V. and Cressoni Araujo, A. (2020) Myokines: A Descriptive Review. The Journal of Sports Medicine and Physical Fitness, 60, 1583-1590. https://doi.org/10.23736/S0022-4707.20.10884-3
[8] Liu, C., Feng, X., Li, Q., Wang, Y., Li, Q. and Hua, M. (2016) Adiponectin, TNF-α and Inflammatory Cytokines and Risk of Type 2 Diabetes: A Systematic Review and Meta-Analysis. Cytokine, 86, 100-109. https://doi.org/10.1016/j.cyto.2016.06.028
[9] Whitham, M. and Febbraio, M.A. (2016) The Ever-Expanding Myokinome: Discovery Challenges and Therapeutic Implications. Nature Reviews Drug Discovery, 15, 719-729. https://doi.org/10.1038/nrd.2016.153
[10] Febbraio, M.A. and Pedersen, B.K. (2020) Who Would Have Thought-Myokines Two Decades on. Nature Reviews Endocrinology, 16, 619-620. https://doi.org/10.1038/s41574-020-00408-7
[11] Lehrskov, L.L. and Christensen, R.H. (2019) The Role of Interleukin-6 in Glucose Homeostasis and Lipid Metabolism. Seminars in Immunopathology, 41, 491-499. https://doi.org/10.1007/s00281-019-00747-2
[12] Fève, B. and Bastard, J.P. (2009) The Role of Interleukins in Insulin Resistance and Type 2 Diabetes Mellitus. Nature Reviews Endocrinology, 5, 305-311. https://doi.org/10.1038/nrendo.2009.62
[13] Kurauti, M.A., Costa-Júnior, J.M., Ferreira, S.M., Santos, G.J., Sponton, C.H.G., Carneiro, E.M., Telles, G.D., Chacon-Mikahil, M.P.T., Cavaglieri, C.R., Rezende, L.F. and Boschero, A.C. (2017) Interleukin-6 Increases the Expression and Activity of Insulin-Degrading Enzyme. Scientific Reports, 7, Article No. 46750. https://doi.org/10.1038/srep46750
[14] Akbari, M. and Hassan-Zadeh, V. (2018) IL-6 Signalling Pathways and the Development of Type 2 Diabetes. Inflammopharmacology, 26, 685-698. https://doi.org/10.1007/s10787-018-0458-0
[15] Ghanem, S.E., Abdel-Samiee, M., Torky, M.H., Gaafar, A., Mohamed, S.M., Salah Eldin, G.M.M., Awad, S.M., Diab, K.A., DM, E.L., Yehia, S.A., Abdelaziz Elbasyouni, H.A. and Elshormilisy, A.A. (2020) Role of Resistin, IL-6 and NH2-Terminal Portion proBNP in the Pathogenesis of Cardiac Disease in Type 2 Diabetes Mellitus. BMJ Open Diabetes Research & Care, 8, e001206. https://doi.org/10.1136/bmjdrc-2020-001206
[16] Adela, R., Reddy, P.N.C., Ghosh, T.S., Aggarwal, S., Yadav, A.K., Das, B. and Banerjee, S.K. (2019) Serum Protein Signature of Coronary Artery Disease in Type 2 Diabetes Mellitus. Journal of Translational Medicine, 17, Article No. 17. https://doi.org/10.1186/s12967-018-1755-5
[17] Moshapa, F.T., Riches-Suman, K. and Palmer, T.M. (2019) Therapeutic Targeting of the Proinflammatory IL-6-JAK/STAT Signalling Pathways Responsible for Vascular Restenosis in Type 2 Diabetes Mellitus. Cardiology Research and Practice, 2019, Article ID: 9846312. https://doi.org/10.1155/2019/9846312
[18] Hong, E.G., Ko, H.J., Cho, Y.R., Kim, H.J., Ma, Z., Yu, T.Y., Friedline, R.H., KurtJones, E., Finberg, R., Fischer, M.A., Granger, E.L., Norbury, C.C., Hauschka, S.D., Philbrick, W.M., Lee, C.G., Elias, J.A. and Kim, J.K. (2009) Interleukin-10 Prevents Diet-Induced Insulin Resistance by Attenuating Macrophage and Cytokine Response in Skeletal Muscle. Diabetes, 58, 2525-2535. https://doi.org/10.2337/db08-1261
[19] Straczkowski, M., Kowalska, I., Nikolajuk, A., Krukowska, A. and Gorska, M. (2005) Plasma Interleukin-10 Concentration Is Positively Related to Insulin Sensitivity in Young Healthy Individuals. Diabetes Care, 28, 2036-2037. https://doi.org/10.2337/diacare.28.8.2036
[20] van Exel, E., J. Gussekloo, A.J. de Craen, M. Frölich, A. Bootsma-Van Der Wiel, and R.G. Westendorp, (2002) Low Production Capacity of Interleukin-10 Associates with the Metabolic Syndrome and Type 2 Diabetes: The Leiden 85-plus Study. Diabetes, 51, 1088-1092. https://doi.org/10.2337/diabetes.51.4.1088
[21] Yssel, H., De Waal Malefyt, R., Roncarolo, M.G., Abrams, J.S., Lahesmaa, R., Spits, H. and de Vries, J.E. (1992) IL-10 Is Produced by Subsets of Human CD4+ T Cell Clones and Peripheral Blood T Cells. The Journal of Immunology, 149, 2378-2384.
[22] Moore, K.W., de Waal Malefyt, R., Coffman, R.L. and O’Garra, A. (2001) Interleukin-10 and the Interleukin-10 Receptor. Annual Review of Immunology, 19, 683-765. https://doi.org/10.1146/annurev.immunol.19.1.683
[23] Yin, Y.W., Hu, A.M., Sun, Q.Q., Zhang, B.B., Liu, H.L., Wang, Q., Zeng, Y.H., Xu,R.J., Zhang, S.J. and Shi, L.B. (2013) Association between Interleukin 10 Gene-1082 A/G Polymorphism and the Risk of Type 2 Diabetes Mellitus: A Meta-Analysis of 4250 Subjects. Cytokine, 62, 226-231. https://doi.org/10.1016/j.cyto.2013.02.025
[24] Forte, G.I., Pilato, G., Vaccarino, L., Sanacore, M., Candore, G., Romano, G.C., Testa, R., Franceschi, C., Capri, M., Marra, M., Bonfigli, A.R., Caruso, C., Scola, L. and Lio, D. (2010) Risk Profiles in Type 2 Diabetes (Metabolic Syndrome): Integration of IL-10 Polymorphisms and Laboratory Parameters to Identify Vascular Damages Related Complications. Current Pharmaceutical Design, 16, 898-903. https://doi.org/10.2174/138161210790883642
[25] Chang, J.S., Chang, C.C., Chien, E., Lin, S.S., Cheng-Shiuan, T., Bai, C.H. and Chao, K.C. (2013) Association between Interleukin 1β and Interleukin 10 Concentrations: A Cross-Sectional Study in Young Adolescents in Taiwan. BMC Pediatrics, 13, Article No. 123. https://doi.org/10.1186/1471-2431-13-123
[26] Zeng, J., Xu, Y., Shi, Y. and Jiang, C. (2018) Inflammation Role in Sensory Neuropathy in Chinese Patients with Diabetes/Prediabetes. Clinical Neurology and Neurosurgery, 166, 136-140. https://doi.org/10.1016/j.clineuro.2018.01.031
[27] Pedersen, B.K., Akerström, T.C., Nielsen, A.R. and Fischer, C.P. (2007) Role of Myokines in Exercise and Metabolism. Journal of Applied Physiology (1985), 103, 1093-1098. https://doi.org/10.1152/japplphysiol.00080.2007
[28] Jabri, B. and Abadie, V. (2015) IL-15 Functions as a Danger Signal to Regulate TissueResident T Cells and Tissue Destruction. Nature Reviews Immunology, 15, 771-783. https://doi.org/10.1038/nri3919
[29] Pérez-López, A., Martin-Rincon, M., Santana, A., Perez-Suarez, I., Dorado, C., Calbet, J.A.L. and Morales-Alamo, D. (2019) Antioxidants Facilitate High-Intensity Exercise IL-15 Expression in Skeletal Muscle. International Journal of Sports Medicine, 40, 16-22. https://doi.org/10.1055/a-0781-2527
[30] Christiansen, T., Bruun, J.M., Paulsen, S.K., Olholm, J., Overgaard, K., Pedersen, S.B. and Richelsen, B. (2013) Acute Exercise Increases Circulating Inflammatory Markers in Overweight and Obese Compared with Lean Subjects. European Journal of Applied Physiology, 113, 1635-1642. https://doi.org/10.1007/s00421-013-2592-0
[31] Wadenpohl, J., Seyfarth, J., Hehenkamp, P., Hoffmann, M., Kummer, S., Reinauer, C., Döing, C., Förtsch, K., Mayatepek, E., Meissner, T. and Jacobsen, M. (2021) CD5-Expressing CD8(+) T-Cell Subsets Differ between Children with Type 1 Diabetes and Controls. Immunology & Cell Biology, 99, 1077-1084. https://doi.org/10.1111/imcb.12488
[32] Stocks, B.T., Wilson, C.S., Marshall, A.F., Hoopes, E.M. and Moore, D.J. (2019) Regulation of Diabetogenic Immunity by IL-15-Activated Regulatory CD8 T Cells in Type 1 Diabetes. The Journal of Immunology, 203, 158-166. https://doi.org/10.4049/jimmunol.1800976
[33] Ge, T., Jhala, G., Fynch, S., Akazawa, S., Litwak, S., Pappas, E.G., Catterall, T., Vakil, I., Long, A.J., Olson, L.M., Krishnamurthy, B., Kay, T.W. and Thomas, H.E. (2020) The JAK1 Selective Inhibitor ABT 317 Blocks Signaling Through Interferon-γ and Common γ Chain Cytokine Receptors to Reverse Autoimmune Diabetes in NOD Mice. Frontiers in Immunology, 11, Article ID: 588543. https://doi.org/10.3389/fimmu.2020.588543
[34] Nadeau, L. and Aguer, C. (2019) Interleukin-15 as a Myokine: Mechanistic Insight into Its Effect on Skeletal Muscle Metabolism. Applied Physiology, Nutrition, and Metabolism, 44, 229-238. https://doi.org/10.1139/apnm-2018-0022
[35] Nagrani, R., Foraita, R., Wolters, M., De Henauw, S., Marild, S., Molnár, D., Moreno, L.A., Russo, P., Tornaritis, M., Veidebaum, T., Ahrens, W. and Marron, M. (2021) Longitudinal Association of Inflammatory Markers with Markers of Glycaemia and Insulin Resistance in European Children. Diabetes/Metabolism Research and Reviews, 38, e3511. https://doi.org/10.1002/dmrr.3511
[36] McPherron, A.C., Lawler, A.M. and Lee, S.J. (1997) Regulation of Skeletal Muscle Mass in Mice by a New TGF-beta Superfamily Member. Nature, 387, 83-90. https://doi.org/10.1038/387083a0
[37] Amor, M., Itariu, B.K., Moreno-Viedma, V., Keindl, M., Jürets, A., Prager, G., Langer, F., Grablowitz, V., Zeyda, M. and Stulnig, T.M. (2019) Serum Myostatin Is Upregulated in Obesity and Correlates with Insulin Resistance in Humans. Experimental and Clinical Endocrinology & Diabetes, 127, 550-556. https://doi.org/10.1055/a-0641-5546
[38] Eilers, W., Cleasby, M. and Foster, K. (2021) Development of Antisense-Mediated Myostatin Knockdown for the Treatment of Insulin Resistance. Scientific Reports, 11, Article No. 1604. https://doi.org/10.1038/s41598-021-81222-7
[39] Chung, J.O., Park, S.Y., Chung, D.J. and Chung, M.Y. (2020) Serum Myostatin Levels Are Positively Associated with Diabetic Retinopathy in Individuals with Type 2 Diabetes Mellitus. Journal of Diabetes and Its Complications, 34, Article ID: 107592. https://doi.org/10.1016/j.jdiacomp.2020.107592
[40] Efthymiadou, A., Vasilakis, I.A., Giannakopoulos, A. and Chrysis, D. (2021) Myostatin Serum Levels in Children with Type 1 Diabetes Mellitus. Hormones (Athens), 20, 777-782. https://doi.org/10.1007/s42000-021-00317-y
[41] Kang, M.J., Moon, J.W., Lee, J.O., Kim, J.H., Jung, E.J., Kim, S.J., Oh, J.Y., Wu, S.W., Lee, P.R., Park, S.H. and Kim, H.S. (2022) Metformin Induces Muscle Atrophy by Transcriptional Regulation of Myostatin via HDAC6 and FoxO3a. Journal of Cachexia, Sarcopenia and Muscle, 13, 605-620. https://doi.org/10.1002/jcsm.12833
[42] Yamakage, H., Tanaka, M., Inoue, T., Odori, S., Kusakabe, T. and Satoh-Asahara, N. (2020) Effects of Dapagliflozin on the Serum Levels of Fibroblast Growth Factor 21 and Myokines and Muscle Mass in Japanese Patients with Type 2 Diabetes: A Randomized, Controlled Trial. Journal of Diabetes Investigation, 11, 653-661. https://doi.org/10.1111/jdi.13179
[43] Kazemi, F. (2019) Myostatin Alters with Exercise Training in Diabetic Rats; Possible Interaction with Glycosylated Hemoglobin and Inflammatory Cytokines. Cytokine, 120, 99-106. https://doi.org/10.1016/j.cyto.2019.04.012
[44] Motahari Rad, M., Bijeh, N., Attarzadeh Hosseini, S.R. and Raouf Saeb, A. (2020) The Effect of Two Concurrent Exercise Modalities on Serum Concentrations of FGF21, Irisin, Follistatin, and Myostatin in Men with Type 2 Diabetes Mellitus. Archives of Physiology and Biochemistry, 12, 1-10. https://doi.org/10.1080/13813455.2020.1829649
[45] Zhang, A., Li, M., Wang, B., Klein, J.D., Price, S.R. and Wang, X.H. (2018) miRNA-23a/27a Attenuates Muscle Atrophy and Renal Fibrosis through Muscle-Kidney Crosstalk. Journal of Cachexia, Sarcopenia and Muscle, 9, 755-770. https://doi.org/10.1002/jcsm.12296
[46] Zhao, B., Han, X., Meng, Q. and Luo, Q. (2018) Early Second Trimester Maternal Serum Markers in the Prediction of Gestational Diabetes Mellitus. Journal of Diabetes Investigation, 9, 967-974. https://doi.org/10.1111/jdi.12798
[47] Li, B., Cui, W. and Yang, J. (2020) Enhanced Skeletal Muscle Growth in MyostatinDeficient Transgenic Pigs Had Improved Glucose Uptake in Stretozotocin-Induced Diabetes. Transgenic Research, 29, 253-261. https://doi.org/10.1007/s11248-020-00194-y
[48] Zubair, M. and Ahmad, J. (2019) Role of Growth Factors and Cytokines in Diabetic Foot Ulcer Healing: A Detailed Review. Reviews in Endocrine and Metabolic Disorders, 20, 207-217. https://doi.org/10.1007/s11154-019-09492-1
[49] Suh, J.M., Jonker, J.W., Ahmadian, M., Goetz, R., Lackey, D., Osborn, O., Huang, Z., Liu, W., Yoshihara, E., van Dijk, T.H., Havinga, R., Fan, W., Yin, Y.Q., Yu, R.T., Liddle, C., Atkins, A.R., Olefsky, J.M., Mohammadi, M., Downes, M. and Evans, R.M. (2014) Endocrinization of FGF1 Produces a Neomorphic and Potent Insulin Sensitizer. Nature, 513, 436-439. https://doi.org/10.1038/nature13540
[50] Kolodziejski, P.A., Sassek. M., Bien. J., Leciejewska. N., Szczepankiewicz. D., Szczepaniak,B., Wojciechowska. M., Nogowski. L., Nowak. K.W., Strowski, M.Z. and PruszynskaOszmalek, E. (2020) FGF-1 Modulates Pancreatic β-Cell Functions/Metabolism: An in Vitro Study. General and Comparative Endocrinology, 294, Article ID: 113498. https://doi.org/10.1016/j.ygcen.2020.113498
[51] Liang, G., Song, L., Chen, Z., Qian, Y., Xie, J., Zhao, L., Lin, Q., Zhu, G., Tan, Y., Li, X., Mohammadi, M. and Huang, Z. (2018) Fibroblast Growth Factor 1 Ameliorates Diabetic Nephropathy by an Anti-Inflammatory Mechanism. Kidney International, 93, 95-109. https://doi.org/10.1016/j.kint.2017.05.013
[52] Ying, L., Wang, L., Guo, K., Hou, Y., Li, N., Wang, S., Liu, X., Zhao, Q., Zhou, J., Zhao, L., Niu, J., Chen, C., Song, L., Hou, S., Kong, L., Li, X., Ren, J., Li, P., Mohammadi, M. and Huang, Z. (2021) Paracrine FGFs Target Skeletal Muscle to Exert Potent Anti-Hyperglycemic Effects. Nature Communications, 12, Article No. 7256. https://doi.org/10.1038/s41467-021-27584-y
[53] Morton, G.J., Matsen, M.E., Bracy, D.P., Meek, T.H., Nguyen, H.T., Stefanovski, D., Bergman, R.N., Wasserman, D.H. and Schwartz, M.W. (2013) FGF19 Action in the Brain Induces Insulin-Independent Glucose Lowering. Journal of Clinical Investigation, 123, 4799-4808. https://doi.org/10.1172/JCI70710
[54] Dolegowska, K., Marchelek-Mysliwiec, M., Nowosiad-Magda, M., Slawinski, M. and Dolegowska, B. (2019) FGF19 Subfamily Members: FGF19 and FGF21. Journal of Physiology and Biochemistry, 75, 229-240. https://doi.org/10.1007/s13105-019-00675-7
[55] Taniguchi, H., Akiyama, N. and Ishihara, K. (2019) Hepatic Fat Content Is Associated with Fasting-Induced Fibroblast Growth Factor 21 Secretion in Mice Fed Soy Proteins. Journal of Nutritional Science and Vitaminology (Tokyo), 65, 515-525. https://doi.org/10.3177/jnsv.65.515
[56] Li, J., Xu, C., Liu, Y., Li, Y., S. Du, Zhang, R., Sun, Y., Zhang, R., Wang, Y., Xue, H.,Ni, S., Asiya, M., Xue, G., Li, Y., Shi, L., Li, D., Pan, Z., Zhang, Y., Wang, Z., Cai, B., Wang, N. and Yang, B. (2020) Fibroblast Growth Factor 21 Inhibited Ischemic Arrhythmias via Targeting miR-143/EGR1 Axis. Basic Research in Cardiology, 115,9. https://doi.org/10.1007/s00395-019-0768-4
[57] Yao, Z., Lin, P., Wang, C., Wang, K. and Sun, Y. (2021) Administration of Metrnl Delays the Onset of Diabetes in Non-Obese Diabetic Mice. Endocrine Journal, 68, 179-188. https://doi.org/10.1507/endocrj.EJ20-0351
[58] Hu, W., Wang, R. and Sun, B. (2021) Meteorin-Like Ameliorates β Cell Function by Inhibiting β Cell Apoptosis of and Promoting β Cell Proliferation via Activating the WNT/β-Catenin Pathway. Frontiers in Pharmacology, 12, Article ID: 627147. https://doi.org/10.3389/fphar.2021.627147
[59] Dadmanesh, M., Aghajani, H., Fadaei, R. and Ghorban, K. (2018) Lower Serum Levels of Meteorin-Like/Subfatin in Patients with Coronary Artery Disease and Type 2 Diabetes Mellitus Are Negatively Associated with Insulin Resistance and Inflam-matory Cytokines. PLOS ONE, 13, e0204180. https://doi.org/10.1371/journal.pone.0204180
[60] El-Ashmawy, H.M., Selim, F.O., Hosny, T.A.M. and Almassry, H.N. (2019) Association of Low Serum Meteorin Like (Metrnl) Concentrations with Worsening of Glucose Tolerance, Impaired Endothelial Function and Atherosclerosis. Diabetes Research and Clinical Practice, 150, 57-63. https://doi.org/10.1016/j.diabres.2019.02.026
[61] Fadaei, R., Dadmanesh, M., Moradi, N., Ahmadi, R., Shokoohi Nahrkhalaji, A., Aghajani, H. and Ghorban, K. (2020) Serum Levels of Subfatin in Patients with Type 2 Diabetes Mellitus and Its Association with Vascular Adhesion Molecules. Archives of Physiology and Biochemistry, 126, 335-340. https://doi.org/10.1080/13813455.2018.1538248
[62] Chung, H.S., Hwang, S.Y., Choi, J.H., Lee, H.J., Kim, N.H., Yoo, H.J., Seo, J.A., Kim, S.G., Kim, N.H., Baik, S.H. and Choi, K.M. (2018) Implications of Circulating Meteorin-Like (Metrnl) Level in Human Subjects with Type 2 Diabetes. Diabetes Research and Clinical Practice, 136, 100-107. https://doi.org/10.1016/j.diabres.2017.11.031
[63] Wu, Q., Dan, Y.L., He, Y.S., Xiang, K., Hu, Y.Q., Zhao, C.N., Zhong, X., Wang, D.G. and Pan, H.F. (2020) Circulating Meteorin-Like Levels in Patients with Type 2 Diabetes Mellitus: A Meta-Analysis. Current Pharmaceutical Design, 26, 5732-5738. https://doi.org/10.2174/1381612826666201007163930
[64] Ali, E.Y., Hegazy, G.A. and Hashem, E.M. (2020) Evaluation of Irisin, Retinol-Binding Protein 4, and Leptin Serum Levels as Biomarkers of Macrovascular Complications Involvement in Saudi Type 2 Diabetes Mellitus. A Case-Control Study. Saudi Medical Journal, 41, 1369-1374. https://doi.org/10.15537/smj.2020.12.25461
[65] Tarboush, N.A., Abu-Yaghi, N.E., Al Ejeilat, L.H., Wahed, R.K.A. and Jeris, I.N.(2021) Association of Irisin Circulating Level with Diabetic Retinopathy: A CaseControl Study. Experimental and Clinical Endocrinology & Diabetes, 129, 36-42. https://doi.org/10.1055/a-0723-3749
[66] Zhu, D., Zhang, X., Wang, F., Ye, Q., Yang, C. and Liu, D. (2022) Irisin Rescues Diabetic Cardiac Microvascular Injury via ERK1/2/Nrf2/HO-1 Mediated Inhibition of Oxidative Stress. Diabetes Research and Clinical Practice, 183, Article ID: 109170. https://doi.org/10.1016/j.diabres.2021.109170
[67] Song, R., Zhao, X., Cao, R., Liang, Y., Zhang, D.Q. and Wang, R. (2021) Irisin Improves Insulin Resistance by Inhibiting Autophagy through the PI3K/Akt Pathway in H9c2 Cells. Gene, 769, Article ID: 145209. https://doi.org/10.1016/j.gene.2020.145209
[68] Duan, H., Ma, B., Ma, X., Wang, H., Ni, Z., Wang, B., Li, X., Jiang, P., Umar, M. and Li, M. (2016) Anti-Diabetic Activity of Recombinant Irisin in STZ-Induced InsulinDeficient Diabetic Mice. International Journal of Biological Macromolecules, 84, 457-463. https://doi.org/10.1016/j.ijbiomac.2015.12.049
[69] Yang, M., Chen, P., Jin, H., Xie, X., Gao, T., Yang, L. and Yu, X. (2014) Circulating Levels of Irisin in Middle-Aged First-Degree Relatives of Type 2 Diabetes Mellitus—Correlation with Pancreatic β-Cell Function. Diabetology & Metabolic Syndrome, 6, Article No. 133. https://doi.org/10.1186/1758-5996-6-133.
This article is excerpted from the Journal of Diabetes Mellitus by Wound World.

